Эволюция
Классическая концепция эволюционной динамики внутри сеӯмейства гоминид, достаточно подробно изложенная в ряде обӯстоятельных книг советских, западноевропейских и американских авторов, базируется на признании нескольких эволюционных стаӯдий, каждая из которых имела достаточно длительную протяженӯность во времени и охватывала несколько локальных форм.
Трудӯно назвать автора, который впервые сформулировал идею стадиӯального подхода, по она, очевидно, интуитивно сознаваласьмногими и медленно, но неуклонно распространялась как теоретиӯческая основа эволюционных взглядов в области антропогенеза. Однако, выделение отдельных стадий имеет своих пионеров, внесӯших наибольший вклад в обоснование стадий как своими фактиӯческими, так и теоретическими разработками. Для рода Pithecan- tropus, обозначаемого еще как совокупность форм архаптропов или протерантропов, это Е. Дюбуа и Ф. Вайденрайх 52, для неӯандертальского вида — это А. Хрдличка 53. Именно им стадиальӯная теория антропогенеза обязана в первую очередь своим распространением и утверждением. И даже те современные исӯследователи, которые выступают, скажем, против неандертальӯской стадии в антропогенезе, защищая гипотезу пресапиенса 54, признают наличие более примитивных морфологических форм и эволюционную динамику внутри семейства в целом.
Нужно, однако, совершенно четко сказать, что концепция эвоӯлюционной стадиальности при всей ее логической четкости, теоӯретической убедительности и фактической аргументированности неоднократно вступала в противоречие с фактическим материаӯлом о локальных вариациях отдельных форм, относящихся к одӯной и той же стадии 55. Это помогло ей избежать метафизичеӯских крайностей, и постепенно она переросла в концепцию внут- ристадиального многообразия, для которой также трудно назвать первого автора и которая интуитивно осознается многими исслеӯдователями.
Автор этих строк полностью разделяет эту концепӯцию, тем более что ее содержание значительно обогатилось мноӯгочисленными блестящими находками последних лет. Поэтому в дальнейшем изложении помимо общей морфологической характеӯристики стадий достаточное внимание будет уделено попыткам морфологического описания и таксономического диагноза отдельӯных локальных форм.Стадия австралопитеков. Морфологическая характеристика этих форм достаточно ясна, тем более что от них найдены кости практически всего скелета 5β. Наиболее богатые местонаӯхождения известны в Южной Африке, хотя теперь ряд находок сделан и в других районах Африки (опи, правда, не описаны пока монографически). Это были, как теперь очевидно, уже полӯностью овладевшие прямохождением формы, что в первую очеӯредь дает все основания рассматривать их как самых ранних из известных пока представителей семейства гоминид. Освобожденӯная от опорной функции рука в дальнейшем еще проделала какую-то эволюцию, так как у самих австралопитековых, как об это можно судить по достаточно фрагментарным материалам, она отличалась значительным разнообразием и нестабильностью строения, а также рядом примитивных особенностей. В строении лицевого скелета отчетливо заметна мегамаксиллярность, сохраӯнившаяся, очевидно, как архаический признак от предыдущей стаӯдии. Усиление этой черты до крайних пределов в некоторых груп-
пах приматов (гигантопитеки, мегантропы) привело, как мы помӯним, к их специализации и вывело их за пределы человеческой линии эволюции. Объем мозга у австралопитеков колебался, по разным определениям, от 500 до 600 см3.
За этим достаточно определенным морфологическим комплекӯсом скрывалось отчетливо выраженное локальное своеобразие. Нет нужды при этом вспоминать только что упомянутых меган- тропов и гигантопитеков. Австралопитековые известны только из Африки, но и там их морфологический полиморфизм очень веӯлик. Уже первые находки 1930—1940-х годов показали, что разӯные формы очень существенно отличаются друг от друга по стеӯпени массивности.
Они были выделены в качестве отдельных родов: грацильные формы в роды Australopithecus и Plesian- thropus, массивные — в род Paranthropus. Однако помимо массивӯности представители этих родов, как об этом свидетельствует их родовой таксономический диагноз, отличались и значительным морфологическим своеобразием. К нему относятся рельеф и форӯма черепа, размеры и рельеф нижних челюстей, некоторые детаӯли строения зубов. Ранний этап развития палеонтологии человеӯка характеризовался преувеличением таксономического значения отдельных форм, и сейчас наблюдается тенденция сократить такӯсономический разрыв между ними, выделяя всего лишь два роӯда: массивных и крупнозубых форм, с одной стороны, грациль- пых — с другой ”. Эта тенденция морфологически абсолютно оправдана, но различия между родами остаются; последнее, кстаӯти сказать, на наш взгляд, лишает значения совмещение всех форм в один род и даже вид. Тенденция такая, ведущая свое начало от таксономической статьи Э. Майра 30-летней давности 58, довольно широко распространилась в американской палеоантроӯпологической литературе ввиду очевидного удобства простых таксономических обозначений (выделяются всего три вида в истории гоминид: Homo transvaalensis, Homo erectus, Homo sapiӯens), но даже разделяющие точку зрения Э. Майра по отношеӯнию к более поздним гоминидам не разделяют ее по отношению к австралопитекам, продолжая выделять внутри них минимум два рода 59.Дополнительный материал доставили раскопки в Восточной Африке, начиная с 1969 г. Если презинджантропа можно без больших сомнений включить в род грацильных австралопитеӯков 60, то зинджантроп очень своеобразен даже по сравнению с массивными австралопитековыми формами 61. Сильным развитиӯем рельефа он напоминал гориллу. Разумеется, его можно расӯсматривать как представителя австралопитековых лишь условно, но объем мозга у этой формы приблизительно соответствует тоӯму, что мы находим у австралопитеков. Другие находки либо представлены отдельными фрагментами скелета, либо столь маӯлыми фрагментами черепов, что их реконструкция остается очень
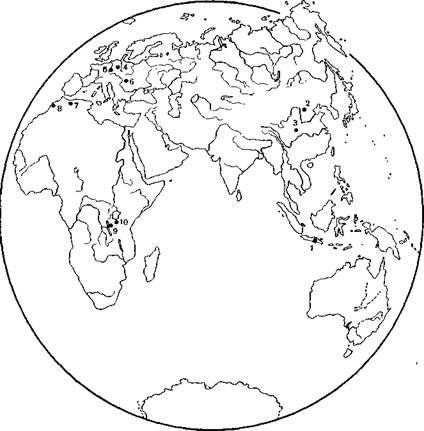
Рис 2 Местонахождения, в которых были обнаружены палеоантропологиӯческие находки, относимые к стадии питекантропов
1 — яванские находки, 2 — Чжоукоудянь, 3 — Ланьтянь, 4 — Бильцингслебен, 5 — Гейдельберг, в — Вертешселлёш, 7 — Тернифин, 8 — Сиди Абдеррахман, 9 — Кооби Фора, 10 — Олдовай
сомнительной.
Но выделение двух родов в составе африканских австралопитеков, по-видимому, объективно, и весьма вероятно, что при широком расселении австралопитековых форм они могли образовывать и другие пока нам неизвестные роды. Таким обраӯзом, высокое таксономическое разнообразие на исходном этапе антропогенеза представляется весьма вероятным.Стадия питекантропов (архантропов, протерантропов). Стадия эта непосредственно следует за стадией австралопитеков, и, по мнению части исследователей, с нее и начинается по-настоящему
развитие семейства гоминид 62. Однако прямохождение и освобоӯждение руки представляются более мощным таксономическим критерием, чем другие морфологические различия между астра- лопитеками и питекантропами, и поэтому нам кажется полностью обоснованным мнение тех исследователей, которые включают первых в семейство Hominidae на правах подсемейства β3. Но это не снимает исключительного значения стадии питекантропов в антропогенезе, так как при переходе к этой стадии и на ее проӯтяжении произошли серьезные морфологические преобразования.
Классические находки питекантропов на Яве и синантропов в Китае выявили огромное сходство этих форм, хотя и обнаруӯжили между ними различия в признаках, демонстрирующих проӯгрессивную эволюцию β4. В первую очередь это касается объема мозга, который у синантропов на 150—200 см3 больше, чем у питекантропов. Все же нет, по-видимому, никаких оснований виӯдеть в них представителей отдельных родов и нужно выделять их, как это и делают многие современные ученые, в качестве отдельных видов. К этому нужно добавить, что выделение солос- ских гоминид в качестве рода, как и объединение их с неандерӯтальцами, вряд ли оправданно. Приведено достаточно много морӯфологических аргументов в пользу того, что наибольшее сходство солосские гоминиды имеют с яванскими питекантропами и, слеӯдовательно, их тоже нужно рассматривать в пределах этой стаӯдии β5.
Новые находки питекантропов на Яве укрепили фактичеӯски базу для морфологической характеристики этой группы, ноӯвые находки на других материках, которые могут быть по тем или иным соображениям поставлены в связь с ней (рис. 2), расӯширили наши представления о ее своеобразии. Его мы коснемӯся позже, а сейчас дадим характеристику представителей этой стадии антропогенеза в целом.Они были среднего или низкого роста и, судя по сохранивӯшимся костям скелета, имели массивный костяк и, следовательӯно, плотное телосложение. Строение костей скелета практически не отличалось от современного, и можно думать, что именно к этой стадии относится полное формирование истинно человечеӯской кисти руки с современными пропорциями. Наличный матеӯриал по кисти (полулунная кость запястья синантропа) дает, правда, этому лишь слабое подтверждение. Череп был в то же время далек от строения черепа современного человека и имел значительное число примитивных особенностей: сильное развитие лицевой части в ущерб мозговой, мощный надбровный и затыӯлочный рельеф, низкий свод и наклонно расположенную лобную кость. Объем внутренней полости черепной коробки колебался вокруг 1000 см3, у питекантропа спускаясь до 900 см3 в средӯнем, у синантропа поднимаясь приблизительно до 1100 см 3, а у солосских гоминид — даже до 1200 см3 (рис. 3). Клювовидная уплощенная форма лобных долей и значительное развитие заты-
лочных долей также являютӯся примитивными признакаӯми в строении мозга у гомиӯнид этой стадии. Но в целом, оценивая строение мозга у них в сравнении с австралоӯпитеками и современным чеӯловеком, можно сказать, что мозг у питекантропов в шиӯроком смысле слова был блиӯже к современному человеӯческому и в качественном и в количественном отношении, чем к мозгу австралопитеков.
Чтобы полностью понять значение новейших находок остатков питекантропов для освещения этой стадии анӯтропогенеза, нужно кратко коснуться их датировки. Еще десятилетие тому назад предӯполагалось, что хронологичеӯский возраст питекантропов Явы не превышает 500— 600 тыс.
лет, возраст синанӯтропов — 300 тыс. лет. По сравнению с австралопитекаӯми, возраст которых уходил вглубь до 1,7-1,8 млн. лет,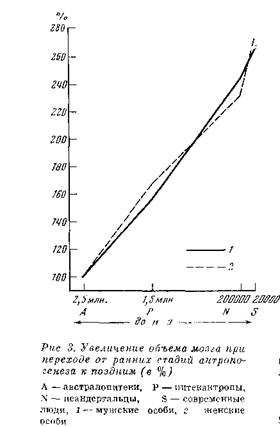
питекантропы справедливо рассматривались как значиӯтельно более молодые формы. Однако последнее десятилетие не только принесло несколько новых находок черепов питекантропов на Яве, расширивших и во многом уточнивших наши знания о морфологии этой группы, но и внесло существенные исправления в их датировку β6.
Новейшие геологические работы на Яве показывают, так же как и исследования фауны из слоев вместе с остатками ранних питекантропов, что датировка их должна быть значительно углубӯлена и восходит ко времени не менее 1,5 млн. лет. Наше предӯставление о датировке синантропов и сейчас остается более или менее прежним и, по-видимому, тем же временем или лишь неӯмногим более ранним могут быть, в свете новейших наблюдений Т. Якоба, датированы и солосские гоминиды. В широких предеӯлах то же можно повторить и про череп, условно считающийся черепом питекантропа и найденный в Восточной Африке в Ол- довае β7. Таким образом, продолжительность рассматриваемой
стадии в антропогенезе приближается к 1,5 млн. лет. Если же относить к группе питекантропов индивидуума, который представӯлен достаточно полно сохранившимся черепом 1470 из формации Кооби Фора (берег оз. Туркане, Восточная Африка) es, для чего, как мы увидим дальше, имеются морфологические основания, то продолжительность стадии питекантропов возрастет еще. Об этом свидетельствуют также и новые находки черепов в Кооби Фора, зафиксированные под номерами 3733 и 3883, морфологиӯчески также близкие к яванским питекантропам и датируемые более чем 1,5 млн. лет 69. Возраст австралопитеков, как показыӯвают новые находки, также удревнился. Весьма вероятно, что обе стадии хронологически перекрывали друг друга и представители австралопитеков достаточно длительное время сосуществовали с типичными представителями стадии питекантропов. Этим фактом стадиальная концепция антропогенеза увязывается с современныӯми представлениями о многолипейной эволюции.
Возможность оценки локального многообразия внутри этой стадии ограничивается значительно большей, чем для предшеӯствующей стадии австралопитеков, фрагментарностью находок, а именно почти полным отсутствием находок черепов с сохранивӯшейся лицевой частью. Только черепа 1470 и 3733 составляют счастливое исключение. Но все же имеются некоторые геологоӯхронологические и морфологические основания для таксономичеӯской дефиниции гоминид этой стадии.
Группа питекантропов Явы, несмотря на некоторое своеобраӯзие отдельных находок, достаточно однородна. Часто писали и своеобразии черепа из Моджокерто, найденного в 1936 г. Р. Кёнигсвальдом и одного из древнейших на Яве. Биологически возраст субъекта из Моджокерто оценивается неодназначно в существующей литературе, по наиболее вероятно считать, что ему было 2—3 года. Объем мозга у взрослой особи был бы примерно 900 см ,, т. е. полностью соответствовал групповой величине. Мноӯгократно отмечавшиеся малые размеры черепа и слабый рельеф также связаны, наверное, с детским возрастом находки. Солос- ские гоминиды отличаются от них гораздо более поздним геолоӯгическим возрастом и отдельными морфологическими чертами, среди которых в первую очередь следует назвать очень большие пазмеры черепа и очень сильное развитие черепного рельефа. Это бесспорно другая самостоятельная группа внутри этой стаӯдии. С. Сартоно, описавший череп питекантропа VIII, объединил его в одну группу с солосскими гоминидами и выделил эту групӯпу в качестве самостоятельного вида внутри рода питекантропов. Морфологическое своеобразие и таксономическая самостоятельӯность этой группы, представленной черепами из Нгапдонга, не вызывают сомнений, но отнесение к этой группе особи питекантӯропа VIII вряд ли можно считать оправданным. С. Сартоно опиӯрался на несколько более крупный объем мозга у особи VIII —
1029 см3. В противоположную, более раннюю группу он объедиӯнил особей 1, II, IV и VII, по его мнению отличавшихся заметно меньшим объемом мозга. Но особь II с объемом мозга в 775 см3, наверное, женская, а череп V, не упомянутый С. Сартоно, имеет объем мозга 975 см3 и заполняет пробел между особью VIII и остальными находками. Все этой заставляет отнестись к гипотезе С. Сартоно с большим сомнением.
Третью группу образуют синантропы, которые, как в солосские гоминиды, хронологически много моложе, чем яванские питеканӯтропы, и, как мы помним, морфологически прогрессивнее их. На права четвертой группы и хронологически, и морфологически, и по своему географическому ареалу могут претендовать те дреӯвнейшие формы этой стадии, которые представлены черепами 1470, 3733 и 3883. Сложнее обстоит дело с олдовайским питеканӯтропом — и по хронологической датировке, и морфологически он близок к солосским формам, но самостоятельный ареал позволяӯет рассматривать его как отдельный вариант. Еще более сложен вопрос о систематической самостоятельности и таксономическом положении таких форм, как атлантроп (как показывает название, выделяемый в качестве рода, что уже было подвергнуто справедӯливой критике 70), гейдельбергский человек 71, находки в Бильӯцингслебен и Вертешсёлёш 72. Последняя имеет огромный объем мозга (1300—1400 см3), но определение объема осуществлено на основе неполного фрагмента затылочной кости и поэтому сомниӯтельно. Гейдельбергская челюсть, как известно, очень примитивӯная, по не могла ли она соединяться с крупной черепной коробӯкой, имеющей относительно крупный внутренний объем? Это не исключено, как и морфологическое сочетание челюстей атлантроӯпа и черепной коробки олдовайского питекантропа. Находка в Бильцингслебен очень фрагментарна. Но разные ареалы в преӯделах Европы и Африки позволяют противопоставлять африканӯские формы европейским и также оценивать их как самостоятельӯные группы. Таким образом, морфологическое разнообразие внуӯтри рассматриваемой стадии антропогенеза может быть сведено к шести отдельным и таксономически самостоятельным группам.
Стадии неандертальцев {палеоантропов). Эта стадия, непоӯсредственно предшествующая появлению человека современного вида и специально выделенная и аргументированная А. Хрдлич- кой73, послужила в истории антропогенеза предметом особенно острой дискуссии. Острота этой дискуссии во многом определяӯлась тем, что с этой стадией в той или иной степени связываӯлась проблема происхождения человека современного вида, сама по себе стоявшая всегда в центре внимания. Гипотеза происхожӯдения человека современного вида не на базе неандертальского вида, а на основе каких-то других форм, принципиально отличавӯшихся от него, имела многих сторонников. Сейчас важно отмеӯтить, что эта гипотеза пе находит подтверждения в имеющихся
морфологических фактах, полученных при изучении единичных находок в Фонтешеваде (Франция) 74, Сванскомбе (Англия) 75 и долине Омо (Эфиопия) 76. Речь идет о находках среднеплейӯстоценового и ранней стадии позднеплейстоценового возраста, которые якобы отличались сапиентными особенностями — отсутӯствием резко выраженного лобного рельефа, высоким черепным сводом и т. д. Тщательное сравнительно-морфологическое изучеӯние этих форм, осуществленное после первых описаний, показало, что первоначальные диагнозы, согласно которым перечисленные находки должны быть отнесены к виду современного человека, не могут быть приняты, так как все находки обнаруживают в дейӯствительности значительное сходство с другими представителями неандертальского вида, а по мнению отдельных исследователей, имеют даже какие-то черты сходства с питекантропами. Наоборот, противоположная гипотеза, основанная па преемственности неӯандертальского и современного видов, опирается на многие морӯфологические факты, не говоря уже о геологических и археологиӯческих, и является достаточно серьезной основой для дальнейӯшего проникновения в проблему, и мы исходим именно из этой гипотезы 77.
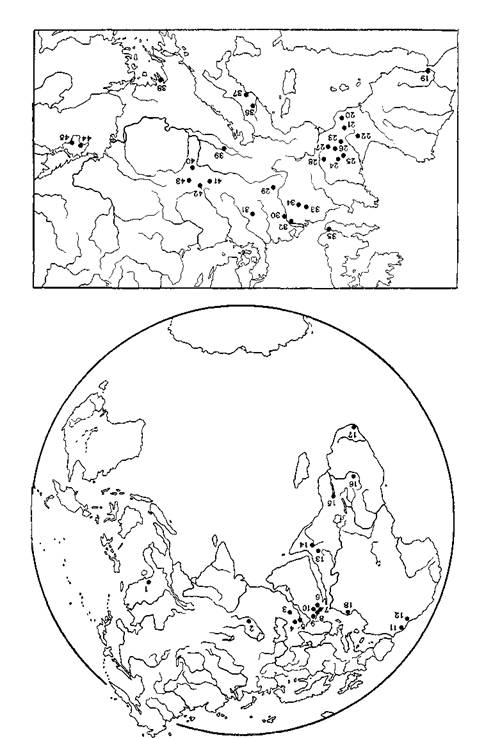
В отличие от предшествующих стадий эта стадия может быть охарактеризована на основании многих находок, достаточно широко распространенных в пространстве (рис. 4). Речь идет о более 500 местонахождениях, из которых происходят костные остатки, допускающие более или менее подробную и ясную морӯфологическую характеристику. Геологическая датировка этих местонахождений колеблется в пределах примерно 40—200 тыс. лет, т. е. эта стадия была несравненно короче, чем предыдуӯщие 7s. Если исходить из среднего индивидуального возраста древних гоминид в 20—25 лет (первая из этих цифр как будто ближе к действительному положению вещей) 79, то на неандерӯтальскую стадию падает 6800—8500 поколений, т. е. на порядок ниже, чем на предыдущие стадии, как об этом уже говорилось. Географически находки неандертальцев охватывают все три материка Старого Света, они неизвестны только в Америке и Австралии, что отражает пространственные пределы тогдашней ойкумены.
Общая морфологическая характеристика неандертальского виӯда может быть сформулирована следующим образом. У подавляӯющего большинства локальных неандертальских форм рост и пропорции тела не изменились по сравнению с представителями предшествующей стадии. Прямохождение было полностью достигӯнуто, и какое-то своеобразие в строении суставных площадок длинных костей конечностей, раньше интерпретировавшееся как доказательство передвижения на полусогнутых нижних конечноӯстях, теперь справедливо рассматривается как функциональное приспособление. Кисть руки имела практически почти современӯ
ное строение, хотя и отличалась, по-видимому, более укороченныӯми пропорциями. Вариациям в строении сустава первой пястной кости, действительно довольно значительным у разных форм, придавалось раньше существенное филогенетическое значение (вплоть до исключения формы, обладавшей уплощенным сустаӯвом, из родословной человека), но теперь очевидно, что приведеӯние большого пальца при неудобной с механической точки зреӯния форме сустава все равно обеспечивалось функционально соответствующей мускулатурой 80.
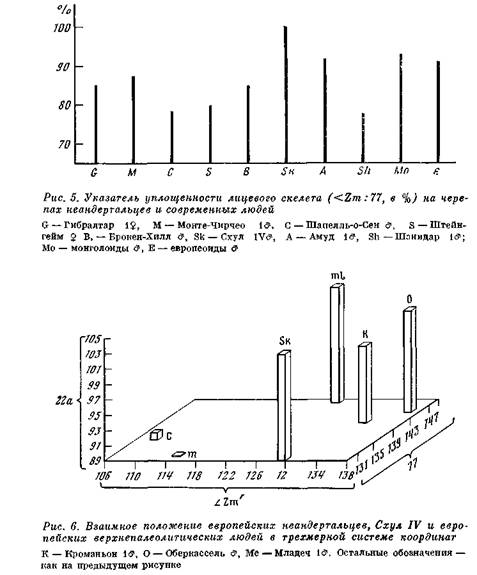
Пожалуй, не будет преувеличением сказать, что наибольшее число примитивных признаков наблюдается у неандертальцев в строении черепа и мозга. Распространенное мнение, что неандерӯтальцы, особенно европейские, отличались каким-то особенно крупным объемом мозга, неверно, так как объем мозга у них все же меньше, чем у верхнепалеолитических людей, приблизительно на 50 см3 и составлял примерно 1450 см3 у мужских особей и 1270 см3 у женских. Клювовидная и уплощенная форма лобных долей, как и значительное развитие затылочной области, преобӯладали у большинства индивидуумов. Во внешнем рельефе череӯпа примитивные особенности проявлялись в переразвитии надӯбровной области, недоразвитии сосцевидных отростков, шиньоноӯобразной форме затылка, уплощенной форме скуловых костей п некоторых других признаках. В частности, структурные элементы горизонтального профиля лицевого скелета находились в ином соӯотношении по сравнению с лицевой морфологией современного человека: при некоторой уплощенности лицевого скелета в верхӯней части он был резко профилирован в нижней (рис. 5) sι. Все это делает неандертальский краниологический комплекс очень своеобразным на фоне других краниологических вариантов древӯних гоминид.
Большое внимание привлек вопрос о локальных вариациях неандертальского типа. Отдельно выделялись африканские форӯмы и яванские формы из Соло 82. Выше говорилось, однако, что последние должны быть включены в группу питекантропов.
Рис.4. Основные местонахождения, в которых были обнаружены палеоантӯропологические находки, относимые к стадии неандертальцев
1 — Мапа, 2 — Тешик-Таш, 3 — Биситун, 4— Тамтама, 5 — Шанидар, в — Схул, 7—Табун, 8— Зуттие, 9 — Амуд, 10 — Кафзех, 11 — Джебель Ирхуд, 12— Рабат, 13 — Рорк Эрик, 14 — Бодо, 15 — Эяси, 1в — Брокен-Хилл, 17 — Салданья, 18 — Хауа- Фтео,
19— Гибралтар, 20 — Баньолас, 21 — Маларно, 22— Монмореи, 21 — Монссмпрон, 24 — Фонтешевад, 25 — Кина, 2в — Ферраси, 27 — Мустье, 28— Шапелль-о-Сен, 29 — Штейнгейм, 30 — Неандерталь, 31 — Эригсдорф, 32 — Энгис, 33 — Спи, 34 — Нолетт, 35 — Сванскомб, Зв — Саккопасторе, 37 — Монте-Чирчео, 38 — Патралона, 39 — Краӯпина, 40 — Шубайюк, 41— Охос, 42— Шипка, 43— Гановцы, 44— Киик-Коба, 45 — Заскальиая
Среди европейских неандертальцев были выделены две группы: более ранняя и морфологически прогрессивная, более поздняя и морфологически примитивная83. Последняя рассматривалась как специализированная форма, либо совсем не принявшая, либо приӯнявшая лишь слабое развитие в формировании человека современного типа. Наконец, на особое место всегда выдвигается
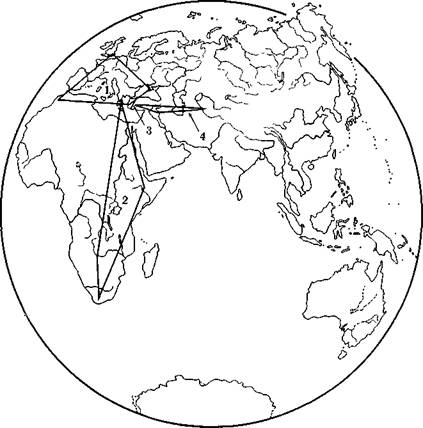
Рис. 7. Схематические ареалы четырех выделенных локальных групп неанӯдертальцев, построенные в соответствии с местоположением соответствуюӯщих находок
1 — европейские неандертальцы, 2 — африканские неандертальцы, 3 — неандертальӯцы группы Схул, 4 — переднеазиатекие неандертальцы
группа Схул (Палестина) как наиболее прогрессивная и явивӯшаяся, по преобладающему мнению, основой формирования соӯвременного человечества 84.
Не касаясь филогенетических взаимоотношений перечисленных локальных групп, что всегда составляет результат большей или меньшей экстраполяции, отметим, что само выделение групп доӯстаточно проблематично. Выделение двух вариантов в составе европейских неандертальцев не находит подтверждения при полӯном анализе метрических и морфологических данных по всем наӯ
ходкам 85. Но туда же, к европейской группе, нужно относить и черепа Схул IV и Табун I, которые практически по всем важӯным признакам не выходят за пределы вариаций в этой группе (рис. 6). Череп неандертальца из Петролоны (Греция) обнаруӯживает значительное число черт, объединяющих его с африканӯскими находками — черепами из Брокен-Хилла и Салданьи86. Самостоятельное место занимает группа Схул, к которой относятӯся индивидуумы Кафзех VI и Зуттие, и самостоятельно также положение таких форм, как Шанидар (Ирак) и Амуд (Палестиӯна), объединяющихся с неандертальцем из Тешик-Таша (Узбеӯкистан). Таким образом, морфологическое разнообразие внутри неандертальской фазы сводится к четырем локальным группам (рис. 7) 87. Особое место должен занимать неандерталец из Мапы (Китай) 88, но его морфологическая характеристика вследӯствие фрагментарности находки неопределенна.
На наш взгляд, в рамках стадии питекантропов выделенные локальные группы вполне могут претендовать на таксономическое положение отдельных видов, тогда как питекантропы в целом выӯделяются как самостоятельный род. Сейчас многие исследователи согласны в том, что неандертальцы образуют один сборный вид и территориальные группы неандертальцев не могут оцениваться иначе, чем локальные расы.
Систематика. Все предшествующее изложение целесообразно подытожить в систематической таблице, полное обоснование котоӯрой было произведено на предшествующих страницах89. Следует подчеркнуть, что при се составлении и обозначении систематичеӯских категорий полностью соблюдено правило приоритета, значеӯние которого неоднократно подчеркивалось в таксономической работе90.
Семейство: человечьи (Hominidae)
1- е подсемейство·, австралопитеки(Subfam. Australopithecinae)
1- й род: австралопитек (Genus Australopithecus)
2- й род: парантроп (Genus Paranthropus)
2- е подсемейство·, люди(Subfam. Homininae)
1- й род: питекантроп (Genus Pithecanthropus)
1- й вид: питекантроп прямоходящий (Species Pithecanthropus erectus)
2- й вид: питекантроп китайский (Species Pithecanthropus pekinensis)
3- й вид: питекантроп солосский (Species Pithecanthropus soloensis)
4- й вид: питекантроп ликский (Species Pithecanthropus leakeyi)
5- й вид: питекантроп кенийский (Species Pithecanthropus kenianensis)
6- й вид: питекантроп гейдельбергский (Species Pithecanthropus heidel- bergensis)
2- й род: человек (Genus Homo)
1-й вид: человек неандертальский (Species Homo neanderthalensis)
1- я локальная раса: человек неандертальский европейский (Homo пеап- deιthalensis europeus)
2- я локальная раса: человек неандертальский африканский (Homo пеап- derthalensis africanus)
3- я локальная раса: человек неандертальский схулский (Homo neander- thalensis skhulensis)
4- я локальная раса: человек неандертальский переднеазиатский (Ното neanderthalensis Asiae occidentali)
2-й вид: человек разумный (Species Homo sapiens)
6.